
コンテンツ
生物学の授業でタンパク質合成について学び始めている人にとって、「転写」と「翻訳」という用語はおそらく馴染みがあるでしょう。これらは、DNA 内のコードが読み取られて特定のタンパク質に翻訳される 2 つの主要な段階です。文字起こしと翻訳、プロセスがどこで行われるか、違いを覚える方法についての短期集中コースが必要な場合は、ここが最適です。

キーポイント
- 転写と翻訳は、遺伝子が読み取られ、対応するタンパク質が作成されるプロセスです。
- 転写により細胞の核内で mRNA が生成されます。
- 翻訳では、mRNA を使用して細胞の細胞質内にタンパク質が生成されます。

背景
、動物、菌類、原生生物、およびほとんどの藻類には
真核細胞が
あります。これは、DNAが存在する核を含む、膜に結合した細胞小器官が含まれていることを意味します。 DNA が核から出ることはありませんが、DNA がコードするタンパク質の生成は核の外側の細胞の細胞質で行われます。これを可能にするためには、転写と翻訳が行われる必要があります。
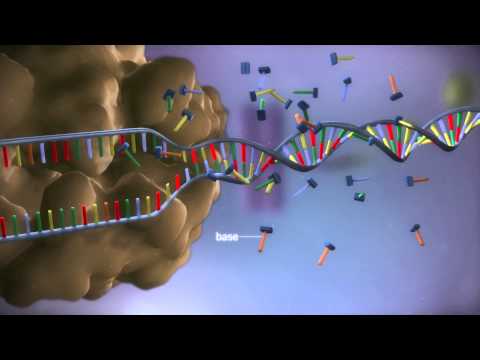
DNA
DNA は複雑な分子です。逆平行の二重らせん構造を持ち、相補的な塩基対が水素結合によって結合されています。言い換えれば、それはねじれたはしごのような二本鎖分子です。はしごの横木はヌクレオチド塩基、つまりアデニン、チミン、グアニン、シトシンです。横木を形成するために、アデニンはチミンと結合し、グアニンはシトシンと結合します。これらのヌクレオチド塩基対は、コドンと呼ばれる 3 つのグループで存在します。コドンは、翻訳中にタンパク質を構築するアミノ酸に対応します。たとえば、DNA の 1 つの鎖が AAT GCG という配列を持っている場合、逆平行鎖上でそれに結合する対応する塩基は TTA CGC の相補的配列を持ちます。
DNA はその複雑な化学構造により、方向性の要素も持っています。単一の DNA 鎖の一方の端は 5′ 端で、もう一方の端は 3′ 端です。相補鎖では、3′ 末端が最初の鎖の 5′ 末端の隣にある逆の向きになります。タンパク質合成の基本を理解するには、転写中に DNA が 3′ から 5′ 方向に読み取られることを知っていれば十分です。論理的な拡張により、転写中に相補的で逆平行なプレ mRNA が 5′ 末端から 3′ 末端に構築されます。
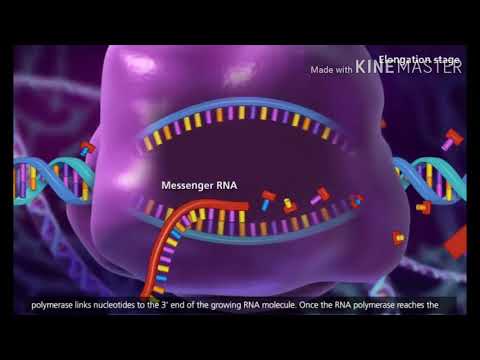
転写: 核内
転写は細胞の核内で起こります。既存の DNA を鋳型として mRNA を書き込むプロセスです。メッセンジャー RNA (mRNA) は、核から細胞の細胞質に移動できる核酸の一種です。 DNA は核から出ることができないため、そのコードは核から出ることができる RNA 分子に転写されます。次の例では、タンパク質 Abc をコードする仮想遺伝子
abc
を想像してください。ここでは、遺伝子
abc
からタンパク質 Abc を合成するための転写の手順を示します。
1) ヘリカーゼが DNA を解く
まず、酵素ヘリカーゼが
abc
遺伝子に対応する DNA 領域を見つけ、塩基間の水素結合を妨げることによってその部分を「巻き戻す」ことになります。ヘリカーゼは DNA を「解凍」し、鋳型鎖と相補鎖 (「コーディング」鎖) を露出させます。鋳型鎖のみが一本鎖プレ mRNA 分子に転写されます。
2) RNA ポリメラーゼはプレ mRNA を合成します
次に、酵素 RNA ポリメラーゼが DNA の鋳型鎖に結合します。 RNA ポリメラーゼは DNA を 3′ 末端から 5′ 末端まで読み取り、適合するヌクレオチドとの結合を形成する反応を触媒します。追加されるヌクレオチドは核内に浮遊していることがわかります。したがって、テンプレート鎖に相補的なヌクレオチドからなるプレ mRNA は、元の
abc
遺伝子のコード鎖と同じ塩基を持ちます。唯一の違いは、プレ mRNA を構築する際にチミンの代わりにヌクレオチドのウラシルが使用されることです。したがって、元の DNA のテンプレート鎖が ATCGAC である場合、コード鎖は TAGCTG である必要があり、pre-mRNA 鎖は UAGCUG になります。
3) 転写後の修飾
abc
遺伝子の pre-mRNA を転写した後、核膜の細孔を通って核外へ出るように準備するための修飾が行われます。プレ mRNA が成熟 mRNA になる前に、3 つの重要な修飾が加えられます。まず、プレ mRNA 分子の 5′ 末端に「5′ キャップ」が付加されます。次に、分子の 3′ 末端に「ポリ (A) テール」が追加されます。最後に、イントロン (RNA 産物では発現されない遺伝子の配列) がプレ mRNA からスプライスされ、エクソン (最終的な RNA で発現される遺伝子の部分) だけが残ります。この記事の目的のために、5′ キャップとポリ (A) テールの化学組成と機能の詳細は必要ありません。
成熟 mRNA の最終構成要素は、5′ キャップ、5′ UTR (開始コドンの前の領域)、エキソン、3′ UTR、および 3′ ポリ(A) テールです。 UTR は、転写後の遺伝子制御に重要な非翻訳領域を指します。 mRNA のこれらのセクションは、開始コドンの前と終止コドンの後の分子の両端に存在します。 5′ および 3′ UTR 配列はタンパク質合成の次の段階では翻訳されず、開始 (開始) コドンと終止コドンの間の配列のみが翻訳されます。

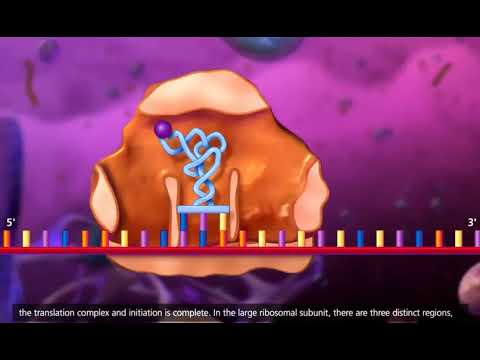
翻訳: 細胞質内
翻訳は、核での転写が終了した後、細胞の細胞質で起こります。これは、リボソームが mRNA 分子をアミノ酸からなるポリペプチド鎖に翻訳するプロセスです。リボソームが翻訳する mRNA の領域は、それぞれがアミノ酸をコードするコドン (3 つのヌクレオチドのセット) で構成されます。 mRNA コドンはアミノ酸に対応し、アミノ酸が結合してポリペプチド鎖を形成し、ポリペプチド鎖が折りたたまれて機能するタンパク質になります。
1) リボソームはポリペプチド鎖を合成します
細胞質では、リボソームの大小のサブユニットが開始コドンで遺伝子
abc
の mRNA 鎖に結合します。開始コドンは常に AUG (アデニン、ウラシル、グアニン) です。細胞質内の tRNA 分子 (転移 RNA) は、リボソームがタンパク質 Abc を形成するポリペプチド鎖を作成するのを助けます。リボソームは mRNA を 5′ 末端から 3′ 末端に向かって読み取ります。一方、tRNA分子は浮遊アミノ酸を対応するコドンに運びます。各 tRNA 分子は各コドンに特異的であり、特定のアミノ酸を伝達します。隣接するアミノ酸間に共有結合が形成され、鎖が形成されます。
2) リボソームがポリペプチド鎖を放出する
リボソームが mRNA を読み取ると、最終的には終止コドンに到達します。アミノ酸に対応しない可能な終止コドンは常に UAA、UAG、または UGA です。これらのコドンを認識する tRNA 分子はなく、放出因子タンパク質がリボソームからの完全なポリペプチド鎖の放出を誘導します。このポリペプチド鎖は折りたたまれて、機能する Abc タンパク質になります。
タンパク質の折り畳み: 細胞質内
翻訳後、ポリペプチド鎖はフォールディングによって複数の構造変換を受けます。最終的なタンパク質の特定の構造がその機能を決定します。この折り畳みは、イオン間の静電力と化学結合の形成によるものです。タンパク質の一次構造はポリペプチド鎖です。それを折りたたむと、タンパク質の二次構造が形成されます。さらに折りたたむと三次構造が形成され、さらに折りたたむと四次構造が形成される場合もあります。 Abc タンパク質が最終形態で機能しているとき、その効果は
abc
遺伝子の発現となります。
転写と翻訳の違いを思い出す
文字起こしと翻訳の順序と基本的なプロセスを覚えておく良い方法は、別の言語で物語を書くことを考えることです。まず、座って、誰かが話すのを聞きながらストーリーを書きます。あなたは物語を書き写します。次に、言語が異なるため、最終製品を作成するために翻訳する必要があります。まずストーリーを書き、それからストーリーを翻訳します。タンパク質合成では、まず mRNA が書き込まれ、次にそれがポリペプチド鎖に翻訳されます。例と同様に、標的遺伝子
abc
が転写され、次に翻訳され、最終的にタンパク質 Abc に折り畳まれます。
転写や翻訳がどこで行われるかを思い出す良い方法は、作用を受ける分子が DNA か mRNA かを思い出すことです。
DNA は決して核から離れることはありません。
したがって、転写中に DNA が作用しているとき、DNA は核内にあるはずであることがわかります。 mRNA が翻訳中に作用するとき、mRNA は細胞質内にあります。
転写と翻訳はどこで行われますか 違いを覚えておくためのヒント イメージギャラリー









関連動画!
Transcription and Translation – Protein Synthesis From DNA – Biology
How to remember Translation and Transcription
")




